DNA鎖の交換反応である相同組換えは体細胞分裂期ではDNA2重鎖切断(Double-strand break; DSB)の修復、減数分裂期では相同染色体の分配に大切な役割を果たしています。
その分子メカニズムは不明な点が多く、我々の研究グループは真核細胞の相同組換えの分子メカニズムあるいは相同組換えの細胞内の機能を明らかにするため以下のような解析を行っています。
体細胞分裂期では相同組換えはDNA障害の修復に重要な役割を果たします。組換えはDNA(染色体)の2重鎖切断で開始し、そのDNA2重鎖末端が削られて生じる1本鎖DNAを利用して、相同な2重鎖DNAを探してきます(下図)。
この反応には大腸菌ではRecA、真核生物ではそのホモログのRad51蛋白質が単鎖DNA上に作る右巻の螺旋構造体(下図)で行なわれると考えられていますが、その詳細については不明です。真核生物ではRad51フィラメントの形成は厳密に制御されていて、さまざまな因子が必要なことが分かっています。例えば、リング上の蛋白質構造体を形成するRad52蛋白質(下図)、最近同定された家族性乳癌の原因遺伝子Brca2もRad51フィラメント形成を助ける補助因子です。我々はRad51のフィラメント形成とその機能を分子レベルで解明することを目指しています。
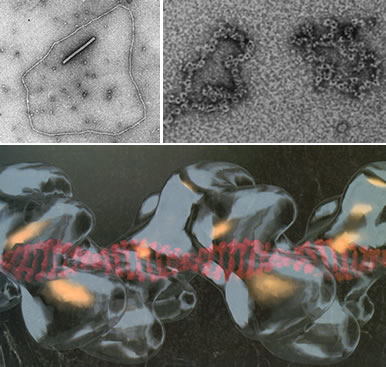
Rad51-DNA(上段左)とその再構築像(下段)、Rad52-DNA(上段右)の電子顕微鏡写真、Rad51はDNA上を右巻きらせん、Rad52はリング構造をとっている。
減数分裂は配偶子形成に必須のプロセスです。減数分裂期ではDNA複製の後、核分裂が2回連続して起こり、中でも第1分裂期では相同染色体が分配される点で非常にユニークです。この相同染色体の間に物理的な結合(キアズマ)を生み出すのが、相同組換えです。減数分裂期の相同組換えは体細胞分裂期の組換えと異なり、交叉型組換えが最終産物として生じるように特殊化しています。
また、染色体あたりの交叉型組換えが起こる数も制御されています。
このように減数分裂期の組換えでは様々な過程が特殊化している訳ですが、中でも前述したDNA鎖間の相同鎖検索、交換反応には体細胞分裂期の組換えに必要なRad51に加えて、減数分裂期特異的なRecAホモログ(Rad51ホモログでもある)Dmc1が必要であることが分かっています(下図)。我々はこの2つのRecAホモログの機能的な協調を明らかにすることが減数分裂期の特異性(交叉型組換えを生み出す原動力)や制御を明らかにするために重要だと考えて、この過程に関わる因子の詳細な解析を行なうことで、減数分裂期の相同鎖検索反応の分子メカニズムを明らかにすることを目指しています。
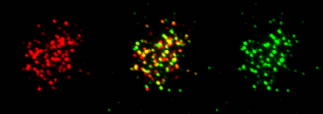
減数分裂期の2つのRecAホモログRad51(緑)、Dmc1(赤)の共局在
染色体の末端構造であるテロメアは染色体の機能の発現には重要な役割を果たしています。
テロメア長は一般にテロメア合成酵素テロメラーゼにより伸長を受け、維持されていますが、テロメラーゼが機能しない時でも細胞はテロメアを維持することができます。
例えば、ある種の癌細胞ではテロメラーゼが発現していないにも関わらず無限に増殖することができます。
このような細胞では相同組換えがテロメラーゼ不在時のテロメアの伸長を保証していると考えられています。
テロメア伸長に関わる組換えには2経路あり、そのうち一方はRecAホモログのRad51に依存していません。
つまり今まで知られていないメカニズム(RecA, Rad51に依存しない形)で相同鎖の検索や交換反応を行なっていると考えられます。パン酵母をモデル系として、このRad51に依存しないテロメアの伸長反応の分子機構の解析を行なっています。
配偶子形成に必要な減数分裂期では第1分裂の前に染色体が核内で配置を変え、動くことが知られています。
この動きは核膜とテロメアの結合や細胞骨格が大切な役割を果たすことが知られていますが、その詳細な運動の仕組みや機能はあまり分かっていません。その分子メカニズムを明らかにすることを目指しています。

減数分裂期の染色体の動き、3秒毎のスナップショットー染色体がダイナミックに動いていることが分かる。
細胞周期S期のDNA複製時の際に内的、外的な要因によってDNA複製フォークがしばしば止まり、複製フォークが崩壊することがしばしば起こります。
複製を完了するためにはこの崩壊した複製フォークを再活性化する必要があります。
大腸菌ではこの活性化に相同組換えが重要な役割を果たすことが知られていますが、真核生物では大腸菌と同じように複製フォークを組換えで再構築しているかどうかすら分かっていません。
ただ、組換えに重要な役割を果たすRad51を欠損した高等真核生物の細胞は生育できないことから、組換えが通常の細胞周期でも必須の機能を持つことは分かっています。
このように真核生物の複製時の組換えを解析するためにはそれを解析するモデル系が必要だと考えられます。
複製フォークの組換えによる再活性化を解明するために複製や組換えについての知見が豊富なパン酵母で解析するためのモデル系を作成しているところです。